


Biologia plantarum 70:47-55, 2026 | DOI: 10.32615/bp.2026.006
Photosynthetic dysfunction and antioxidant defense disruption in rice seedlings induced by tire wear microplastics
- School of Life and Health Sciences, Hainan Province Key Laboratory of One Health, Collaborative Innovation Center of Life and Health, Hainan University, Haikou, Hainan 570228, P.R. China
Background and aims: Tire wear microplastics (TWMs) are emerging environmental contaminants, but their ecological risks to agricultural systems remain poorly understood.
Methods: Rice seedlings were exposed to 0, 10, 100, and 1 000 mg L-1 TWMs for 10 days. Growth parameters, chlorophyll fluorescence, and antioxidant enzyme activities were measured.
Results: TWMs promoted growth; exposure to 1 000 mg L-1 TWMs increased plant height and root length by 8.06% and 57.38%, respectively. Chlorophyll fluorescence analysis revealed that TWMs significantly suppressed rETRmax by 32.53 - 43.62% and altered qP and NPQ. TWMs inhibited Y(NPQ) while enhancing Y(NO) loss, indicating impaired photoprotective dissipation and aggravated photodamage. TWMs also inhibited SOD, POD, CAT, and APX activities in both leaves and roots, with root CAT and APX decreasing by up to 37.35% and 40.34%, reflecting a direct impairment of the antioxidant defense system.
Conclusions: Rice seedlings achieve TWM-induced short-term growth at the expense of compromised photosynthetic efficiency and antioxidant defense, leading to an unsustainable compensatory state. This study provides physiological evidence for assessing TWMs phytotoxicity in agricultural systems.
Keywords: antioxidant system, chlorophyll fluorescence, photosynthesis, rice, tire wear microplastics.
Received: December 15, 2025; Revised: March 6, 2026; Accepted: March 20, 2026; Published online: April 7, 2026 Show citation
| ACS | AIP | APA | ASA | Harvard | Chicago | Chicago Notes | IEEE | ISO690 | MLA | NLM | Turabian | Vancouver |
Introduction
Global tire production continues to rise markedly, driven by the expansion worldwide vehicle fleet. For instance, China’s tire output alone reached 449 million units in the first half of 2021 (Osman et al., 2025), leading to substantial emissions of tire wear microplastics (TWMs). TWMs are continuously generated by the mechanical abrasion of tires against road surfaces. Owing to their persistence and high environmental mobility, TWMs have been detected in diverse environmental compartments, including the atmosphere, pedosphere, surface water, and sediments (Wang et al., 2023a), posing potential risk to ecosystems and human health. A synthesis of available research indicates that TWMs are a dominant fraction of environmental microplastics, with concentrations in road dust reaching 1.7 - 11.3 g kg-1 (Goßmann et al., 2021), exceed total microplastic loads in runoff and sediments by 13- to 49-fold (Eisentraut et al., 2018); and predicted maximum levels in surface waters ranging from 0.03 to 56 mg L-1 (Wik and Dave, 2009).
The widespread environmental presence of TWMs has raised considerable concern about their ecotoxicological risks. Evidence suggests that TWMs adversely affect organisms through three principal mechanisms: physical damage, chemical toxicity, and combined effects (Campanale et al., 2022). Physical damage occurs following the primary exposure routes-inhalation, dermal contact, and ingestion (Luo et al., 2021). Internalized particles can translocate and induce direct physical interference, such as impairing digestive function in the gastrointestinal tract (Gopinath et al., 2019). Concurrently, exposure to TWMs can trigger cytoskeletal rearrangement in pulmonary epithelial cells, a mechanism linked to fibrotic lung damage (Li et al., 2022). In soil fauna, TWMs ingestion further triggers dysbiosis of the gut microbiota (Ding et al., 2020). In terms of chemical toxicity and combined effects, TWMs leachates, containing synthetic rubber monomers, additives (e.g., antioxidants and plasticizers), and adsorbed heavy metals, such as zinc, elicit oxidative stress, neurotoxicity, and endocrine disruption (Wik and Dave, 2009). Moreover, TWMs can act as a vectors for co-contaminants, potentially amplifying overall toxicity (Sathicq et al., 2022).
While concerns over microplastic pollution are mounting, the specific effects of TWMs on plants are poorly understood. Agricultural soils are a major sink for microplastics from various sources, with TWMs constituting a significant but often underestimated input (Galafassi et al., 2019). Existing research on MPs reveals multifaceted impacts, with a critical initial step being physical penetration – MPs enter through root cracks and stomata, then translocate within the vascular system via the transpiration stream (Li et al., 2020a; Lian et al., 2021). Upon entry into plant tissues, MPs trigger a cascade of physiological toxicities, typically manifested as growth inhibition, including reduced biomass, plant dwarfing, and impaired leaf development (Li et al., 2020b). Simultaneously, studies have shown that TWMs can inhibit root elongation, photosynthetic efficiency, and carbohydrate metabolism in the aquatic plant Lemna minor (Putar et al., 2025). Research on the effects of TWMs on food crops remains scarce. Beyond aquatic species, recent studies have also revealed species-specific responses to TWMs in terrestrial crops. Wasnik et al. (2026) reported that mung bean and tomato exhibited distinct responses to TWMs leachates: tomato relied on high SOD activity in leaves as its primary defense, whereas mung bean showed reduced SOD activity under high TWMs leachate concentrations, leading to uncontrolled ROS accumulation and subsequent lipid peroxidation. These species-specific differences highlight the complexity of TWMs phytotoxicity and underscore the need for systematic investigations across diverse crop species, particularly staple crops such as rice. For a 10-day exposure, we focused on early responsive indicators: root length and shoot height as rapid morphological traits (root elongation being particularly sensitive due to direct contact with particles), together with photosynthetic parameters and antioxidant enzyme activities as physiological endpoints. Physiological parameters such as photosynthetic performance and antioxidant enzyme activities are widely recognized as early-warning indicators of plant stress, often preceding changes in growth endpoints such as biomass accumulation (Ralph and Gademann, 2005; Zuo et al., 2025). This combination enables detection of initial stress responses within a short timeframe, providing a comprehensive assessment of early TWMs effects on rice seedlings.
Given the potential yet uncertain risks that TWMs pose to staple crops, this study utilized rice (Oryza sativa L.) as a model system to systematically evaluate the effects of micron-sized (100 µm) TWMs across a range of concentrations on growth, leaf photosynthetic parameters, and antioxidant enzyme activities. The objective was to elucidate the effects of TWMs, as an emerging environmental pollutant, on crop growth and physiological indicators, thereby providing critical insights for a more comprehensive risk assessment of its ecological impacts in agricultural systems.
Materials and methods
Experimental material and treatments: TWMs with a mean particle size of 100 µm were purchased from Dujiangyan Huayi Rubber Co. This specified size falls within the 20 - 212 µm range representative of TWMs commonly found in road dust (Chae et al., 2024), ensuring environmental relevance. Seeds of the japonica rice (Oryza sativa L.) cultivar 'Zhonghua 11' were used. Uniform and plump rice seeds were surface-sterilized by soaking in 0.1% H2O2 solution for 6 h, followed by 5 - 6 rinses with deionized water. The sterilized seeds were then immersed in deionized water in a 50-mL centrifuge tube and incubated in darkness at 30°C for 36 h for imbibition. Subsequently, the seeds were transferred to Petri dishes containing moist filter paper and kept in the dark at 30°C for 3 d to germinate. Germinated seedlings were transferred to a controlled climate chamber set at 25°C. Uniform seedlings were selected and transplanted into a six-cell hydroponic container (12.7 cm × 8.7 cm × 8.7 cm × 11.4 cm) containing Kimura B nutrient solution, with 12 seedlings per container. Three replicate containers were established for each treatment, resulting in a total of 36 seedlings per treatment. The plants were grown under controlled climate chamber: 25°C, a photosynthetic photon flux density of 160 µmol m-2 s-1, and a 16/8-h light/dark photoperiod. After a 16-day preculture, short-term treatments were initiated by amending the nutrient solution with TWMs at concentrations of 0 (control), 10, 100, and 1 000 mg L-1. A 10-day exposure period was selected to investigate the early physiological responses of rice seedlings to TWMs, as previous studies have demonstrated that microplastic-induced physiological changes in plants can be detected within 10 d of exposure (Liu et al., 2025). After 10 d of TWMs exposure, plant growth and physiological parameters were assessed.
Growth parameters: To ensure initial uniformity, shoot height and root length were measured immediately before initiating TWMs exposure and again at the end of the 10-day exposure period. For each measurement time point, these parameters were assessed on the representative seedlings from each treatment group.
Chlorophyll fluorescence: After 10 d of treatment, chlorophyll fluorescence parameters of photosystem II (PSII) were measured on the rice seedlings. Prior to measurement, the seedlings were dark-adapted for 20 min. Measurements were conducted on fully expanded leaves using a pulse-amplitude modulated fluorometer (PAM-2500, Walz, Germany). The rapid light curves (RLCs) were conducted by exposing the samples to light intensities at different gradients (0, 20, 84, 237, 449, 811 µmol photons m-2 s-1) at approximately 2 cm from the leaf tip. The RLCs were fitted to obtain the maximum relative electron transport rate (rETRmax) and the effective quantum yield of PSII [Y(II)]. Furthermore, the key chlorophyll fluorescence parameters included photochemical fluorescence quenching (qP), non-photochemical fluorescence quenching (NPQ), light-induced non-photochemical fluorescence quenching [Y(NPQ)], and non-light-induced non-photochemical fluorescence quenching [Y(NO)] were recorded.
Soluble protein and antioxidant enzyme activities: Leaf (0.05 g) and root (0.2 g) samples were flash-frozen in liquid nitrogen and homogenized in pre-chilled mortars with 3 mL (2 mL for roots) of phosphate buffer (PBS; 0.05 M, pH 7.8). The homogenates were centrifuged at 5 500 × g for 10 min at 4°C. The resulting supernatant was used for subsequent analyses. Soluble protein (SP) content in the supernatant was quantified according to the Bradford method (Bradford, 1976). Briefly, 10 µL of supernatant was mixed with 210 µL of Coomassie Brilliant Blue G-250 reagent in a microplate well. The absorbance at 595 nm was measured using a microplate reader. Protein concentration was calculated from the standard curve prepared with bovine serum albumin. The remaining supernatant was used for subsequent antioxidant enzyme activity analyses. Superoxide dismutase (SOD) activity was determined by monitoring the inhibition of nitroblue tetrazolium (NBT) photoreduction at 560 nm (Batool et al., 2020). The reaction mixture (240 µL) contained 50 mM PBS (pH 7.8), 130 mM methionine, 750 µM NBT, 100 µM EDTA, 20 µM riboflavin, and 20 µL enzyme extract. Peroxidase (POD) activity was assayed by measuring the oxidation of guaiacol at 470 nm (Kavian et al., 2022). The reaction mixture (270 µL) contained 5.4 mM guaiacol, 29.4 mM H2O2, and 90 µL enzyme extract. Catalase (CAT) activity was measured by monitoring the decomposition of H2O2 at 240 nm (Goswami and Das, 2016). The reaction mixture (3 mL) contained 1.9 mL of distilled water, 1 mL of 29.4 mM H2O2 prepared in 50 mM PBS (pH 7.0), and 0.1 mL enzyme extract. Ascorbate peroxidase (APX) activity was evaluated by monitoring the decrease in absorbance at 290 nm due to ascorbate oxidation (Batool et al., 2020). The reaction mixture (3 mL) contained 50 mM PBS (pH 7.0), 15 mM ascorbate, 15 mM H2O2, and 200 µL enzyme extract.
Statistical analysis: All data were obtained from three independent biological replicates per treatment (n = 3), with each replicate consisting of 12 seedlings pooled from one hydroponic container. Data organization was performed using Microsoft Excel 2019. Statistical analyses were conducted with SPSS 23.0 (SPSS Inc., Chicago, IL, USA) employing one-way analysis of variance (ANOVA) followed by Duncan's multiple range test to determine significant differences among treatment groups at a significance level of P < 0.05. Specifically, the parameters including RLCs, qP, NPQ, Y(NPQ), and Y(NO) were analyzed using a mixed linear model in R software version 4.2.2 for Windows (R Foundation for Statistical Computing, Vienna, Austria). All figures were prepared using GraphPad Prism 8.0.2 (GraphPad Software, San Diego, CA, USA).
Results
Effects of TWMs on rice seedling growth parameters: No significant differences in shoot height or root length were detected among the treatment groups prior to TWMs exposure (Fig. 1A), confirming initial uniformity of the seedlings. After 10 d of exposure, statistical analysis revealed a treatment-dependent growth promotion by TWMs. Specifically, shoot height increased by 8.06% in the 1 000 mg L-1 treatment group compared to the control. A more pronounced stimulatory effect was observed on root length, which exhibited a maximum increase of 57.38% at 1 000 mg L-1 TWMs (Fig. 1B).
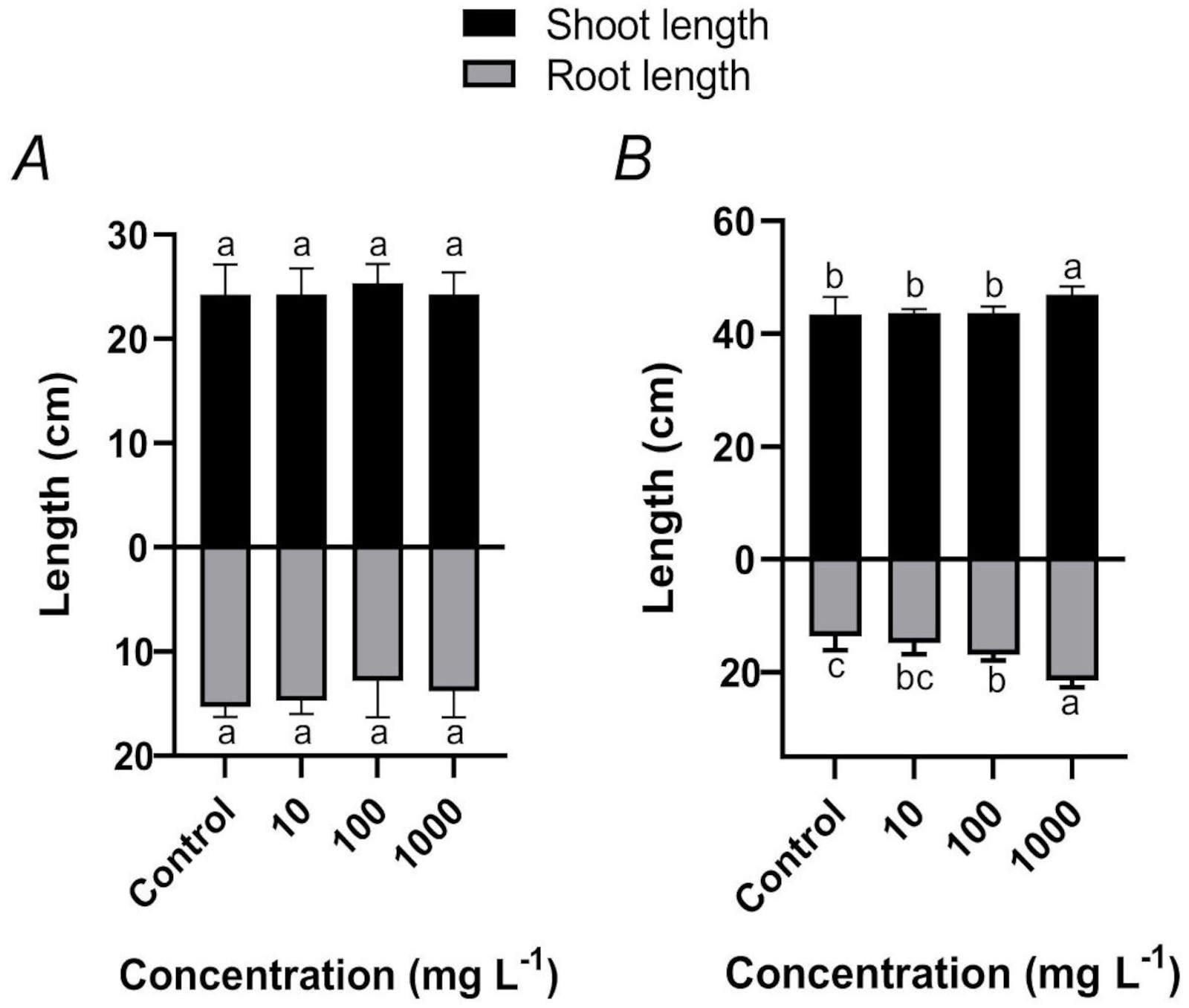
Fig. 1. Effects of TWMs on the growth of rice seedlings. (A) Shoot height and root length of seedlings at the start of the experiment (day 0). (B) Shoot height and root length of seedlings after 10 d of exposure to TWMs (10, 100, and 1 000 mg L-1). Data are represented as mean ± SD (n ≥ 3). Different letters indicate significant differences among treatments (P < 0.05, Duncan's multiple test).
Effects of TWMs on the photosynthetic performance of PSII in rice seedlings: The Y(II) in rice seedlings showed no statistically significant differences following exposure to TWMs compared to the control group (Fig. 2A), indicating maintained baseline efficiency of light energy conversion in PSII reaction centers. In contrast, rETRmax was significantly suppressed across all TWMs treatments, with inhibition rates of 32.53, 43.62, and 40.68% at concentrations of 10, 100, and 1 000 mg L-1, respectively (Fig. 2B). Although the 100 mg L-1 TWMs treatment resulted in the strongest suppression, no statistically significant differences were detected among the different concentration groups. Analysis of RLCs revealed that rETR increased linearly with PAR at low light intensities (0 - 200 μmol photons m-2 s-1). At higher PAR levels (300 - 800 μmol photons m-2 s-1), the rETR growth rate slowed, approaching saturation. Notably, the inhibitory effect of TWMs exposure on electron transport became more pronounced with increasing TWMs concentration as PAR exceeding 300 μmol photons m-2 s-1. Statistical analysis confirmed that all TWM-treated groups showed significantly suppressed RLCs compared to the control, with the degree of suppression exhibiting a clear concentration-dependent pattern (Fig. 2C).
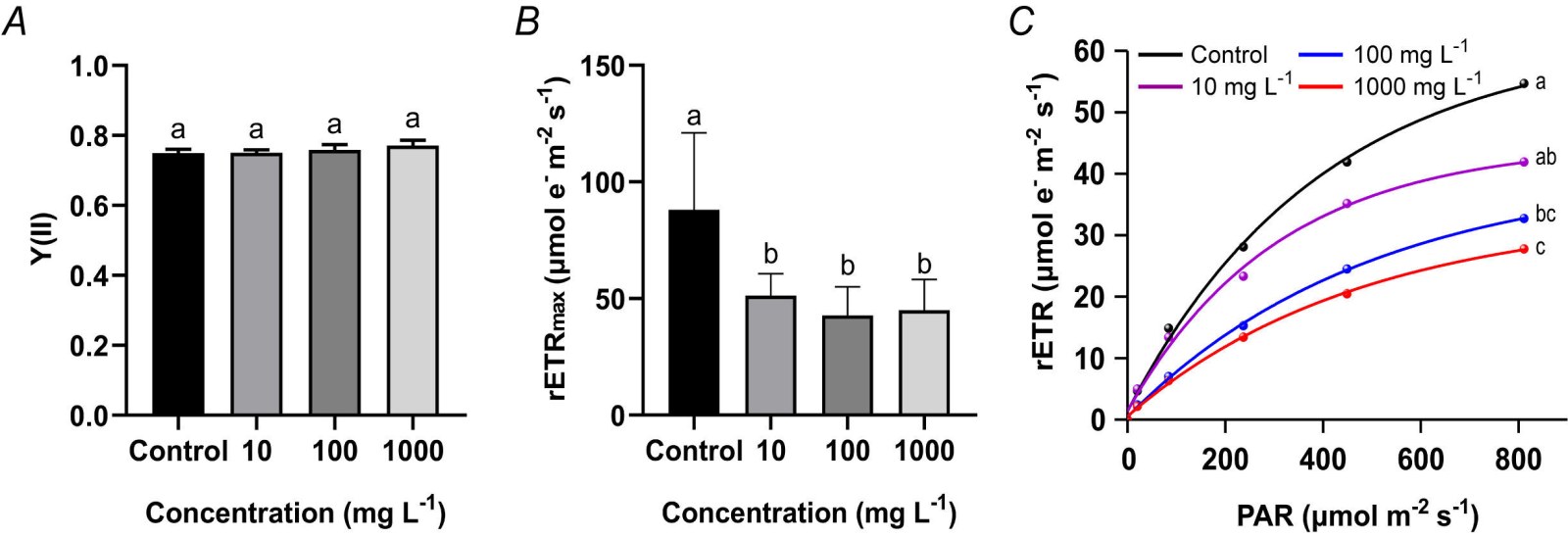
Fig. 2. Effect of a 10-day exposure to TWMs (10, 100, and 1 000 mg L-1) on the photosynthetic parameters of rice seedlings. (A) Effective quantum yield of PSII [Y(II)]. (B) Maximum relative electron transport rate (rETRmax). (C) Rapid light curves (RLCs). Data are represented as mean ± SD (n = 3). Different letters indicate significant differences between the treatments (P < 0.05, Duncan's multiple test).
Effects of TWMs on key chlorophyll fluorescence parameters in rice seedlings: qP decreased with increasing PAR in all treatment groups. The 1 000 mg L-1 TWMs exposure showed the most attenuated decline, while the 10 mg L-1 exposure exhibited a relatively steeper decrease (Fig. 3A). Conversely, NPQ increased with PAR intensity, with the final highest NPQ value in the control group (1.132). All the TWM-treated groups (10, 100, and 1 000 mg L-1) reached significantly lower values of 0.661, 0.767, and 0.750, respectively, indicating concentration-dependent suppression of NPQ induction (Fig. 3B). Y(NPQ) exhibited a consistent upward trend across all experimental groups with increasing PAR levels. However, the magnitude of this increase was consistently attenuated in all TWM-treated seedlings compared to the control, with the 100 mg L-1 TWM-treatment group showing a significant reduction in the extent of Y(NPQ) rise (Fig. 3C). As illustrated in Fig. 3D, Y(NO) values were comparable among all groups under low PAR intensities. As PAR levels increased, Y(NO) exhibited a characteristic pattern of an initial increase followed by a subsequent decline. Notably, the extent of the decrease was markedly less pronounced in all TWM-treated groups relative to the control.
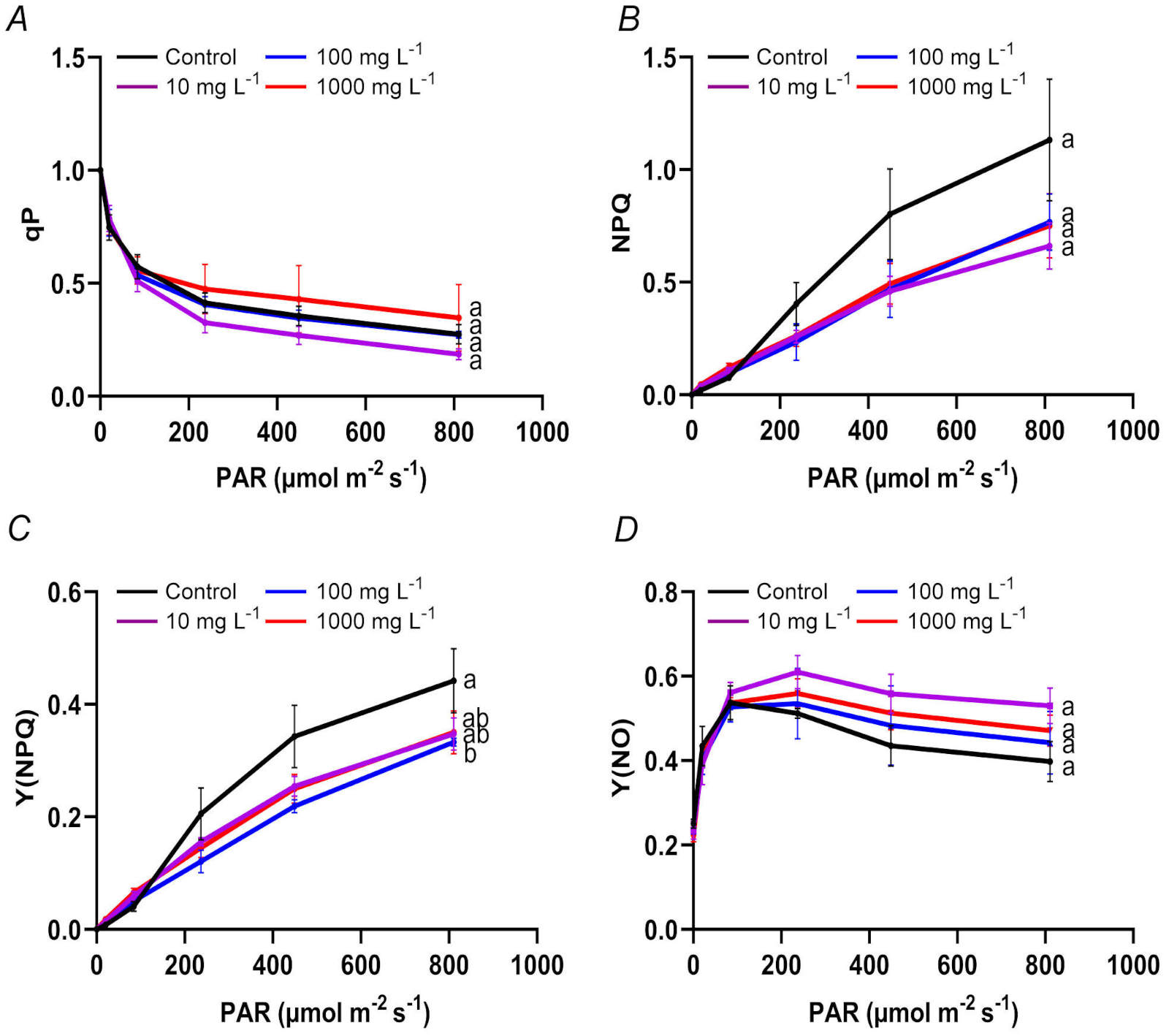
Fig. 3. Effects of a 10-day exposure to TWMs (10, 100, and 1 000 mg L-1) on chlorophyll fluorescence parameters in rice seedlings. (A) Photochemical fluorescence quenching (qP). (B) Non-photochemical fluorescence quenching (NPQ). (C) Light-induced non-photochemical fluorescence quenching [Y(NPQ)]. (D) Non-light-induced non-photochemical fluorescence quenching [Y(NO)]. Data are represented as mean ± SD (n = 3). Different letters indicate significant differences between the treatments (P < 0.05, Duncan's multiple test).
Effects of TWMs on soluble protein content in rice seedlings: TWMs exposure significantly influenced the accumulation of SP in rice seedlings, with distinct response patterns observed between leaves and roots. In leaves, SP content significantly increased by 32.50, 35.39, and 28.75% under 10, 100, and 1 000 mg L-1 TWMs treatments, respectively, compared to the control. The most pronounced enhancement was observed at 100 mg L-1 (Fig. 4A). In roots, SP content also increased in a concentration-dependent manner, rising by 6.91, 32.50, and 11.63% at the respective concentrations, with the peak level similarly occurring at 100 mg L-1 (Fig. 4B).
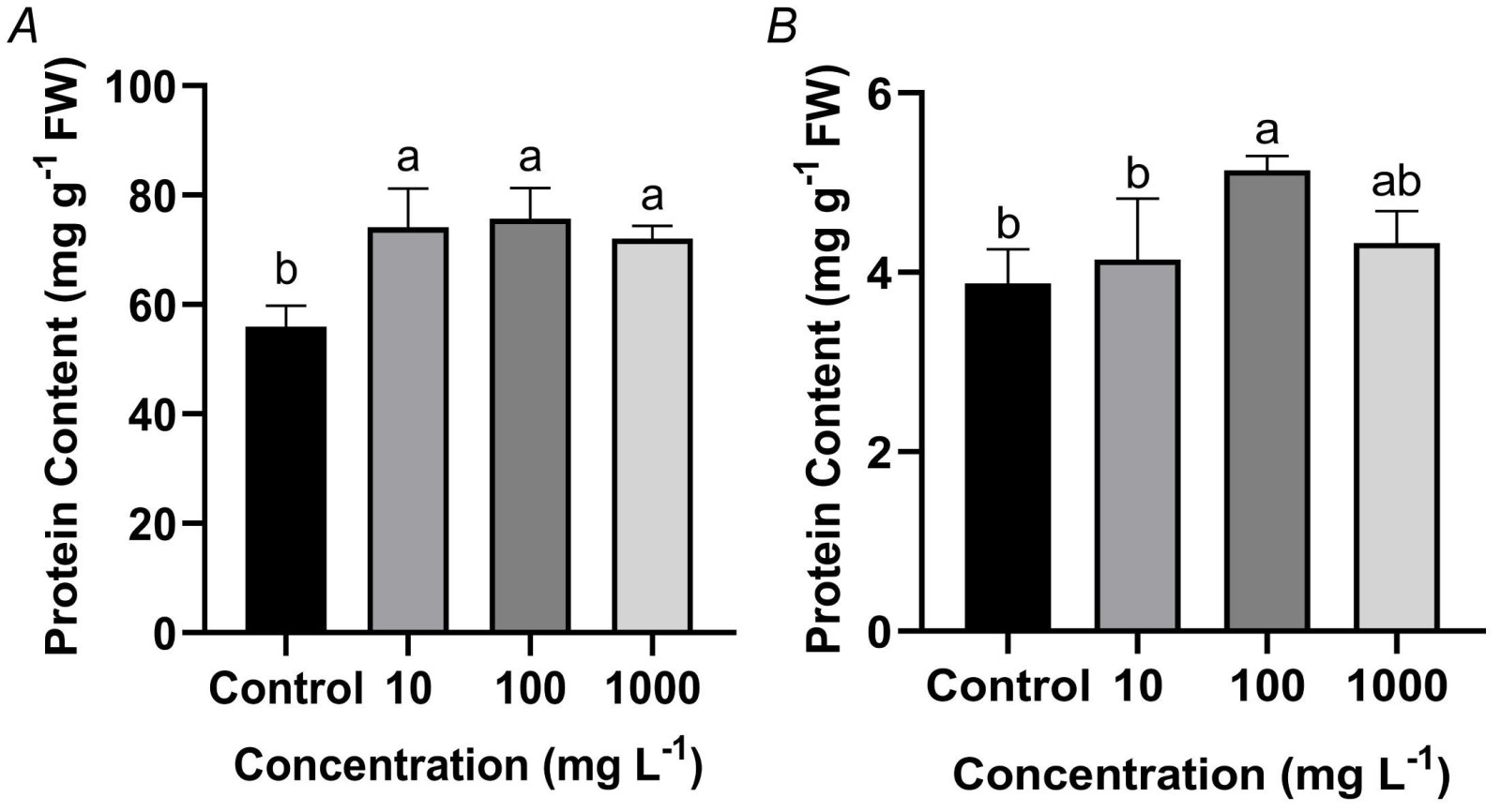
Fig. 4. Effect of a 10-day exposure to TWMs (10, 100, and 1 000 mg L-1) on soluble protein (SP) content in rice seedlings. (A) SP content in leaves. (B) SP content in roots. Data are represented as mean ± SD (n = 3). Different letters indicate significant differences between the treatments (P < 0.05, Duncan's multiple test). FW - fresh weight.
Effects of TWMs on antioxidant enzyme activities in rice seedlings: To investigate the oxidative stress response, the activities of key antioxidant enzymes (SOD, POD, CAT, and APX) were measured in leaves and roots of rice seedlings under TWMs exposure. In leaves, contrary to a typical activation of defense, the results revealed a general suppression of the antioxidant system. SOD activity was significantly inhibited across all treatment groups compared to the control (Fig. 5A). POD activity also exhibited a decreasing trend, with a significant suppression at 10 mg L-1 TWMs (Fig. 5B). However, no statistically significant differences in the activities of CAT and APX were detected between the TWM-treated groups and the control (Fig. 5C,D).
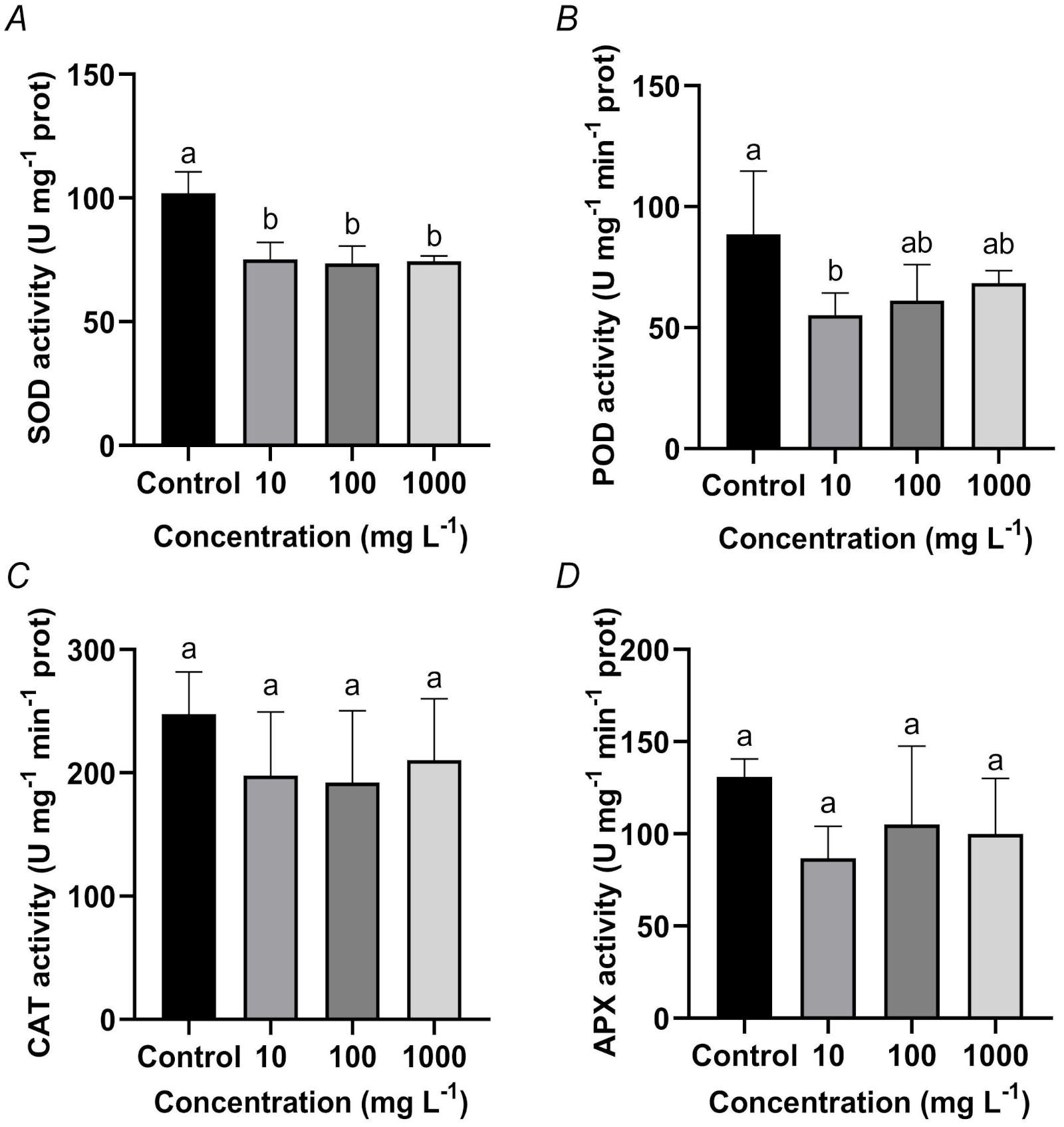
Fig. 5. Effect of a 10-day TWMs (10, 100, and 1 000 mg L-1) treatment on the activities of leaf antioxidant enzymes in rice seedlings. (A) Superoxide dismutase (SOD). (B) Peroxidase (POD). (C) Catalase (CAT). (D) Ascorbate peroxidase (APX). Data are represented as mean ± SD (n = 3). Different letters indicate significant differences between the treatments (P < 0.05, Duncan's multiple test).
The activities of antioxidant enzymes in roots displayed distinct responses to TWMs exposure. SOD activity showed a concentration-dependent inhibition, with a significant suppression observed at 100 mg L-1 TWMs, while the effects at 10 and 1 000 mg L-1 did not reach statistical significance (Fig. 6A). POD activity remained unchanged across all treatment groups (Fig. 6B). In contrast, both CAT and APX activities exhibited clear concentration-dependent suppression. Both enzymes activities progressively decreased with increasing TWMs concentration. Specifically, CAT activity was declined by 9.49, 34.96, and 37.35% at 10, 100, and 1 000 mg L-1 TWMs, respectively, compared to the control (Fig. 6C). Similarly, APX activity was reduced by 25.95, 27.30, and 40.34% in the TWM-treated groups relative to the control (Fig. 6D).
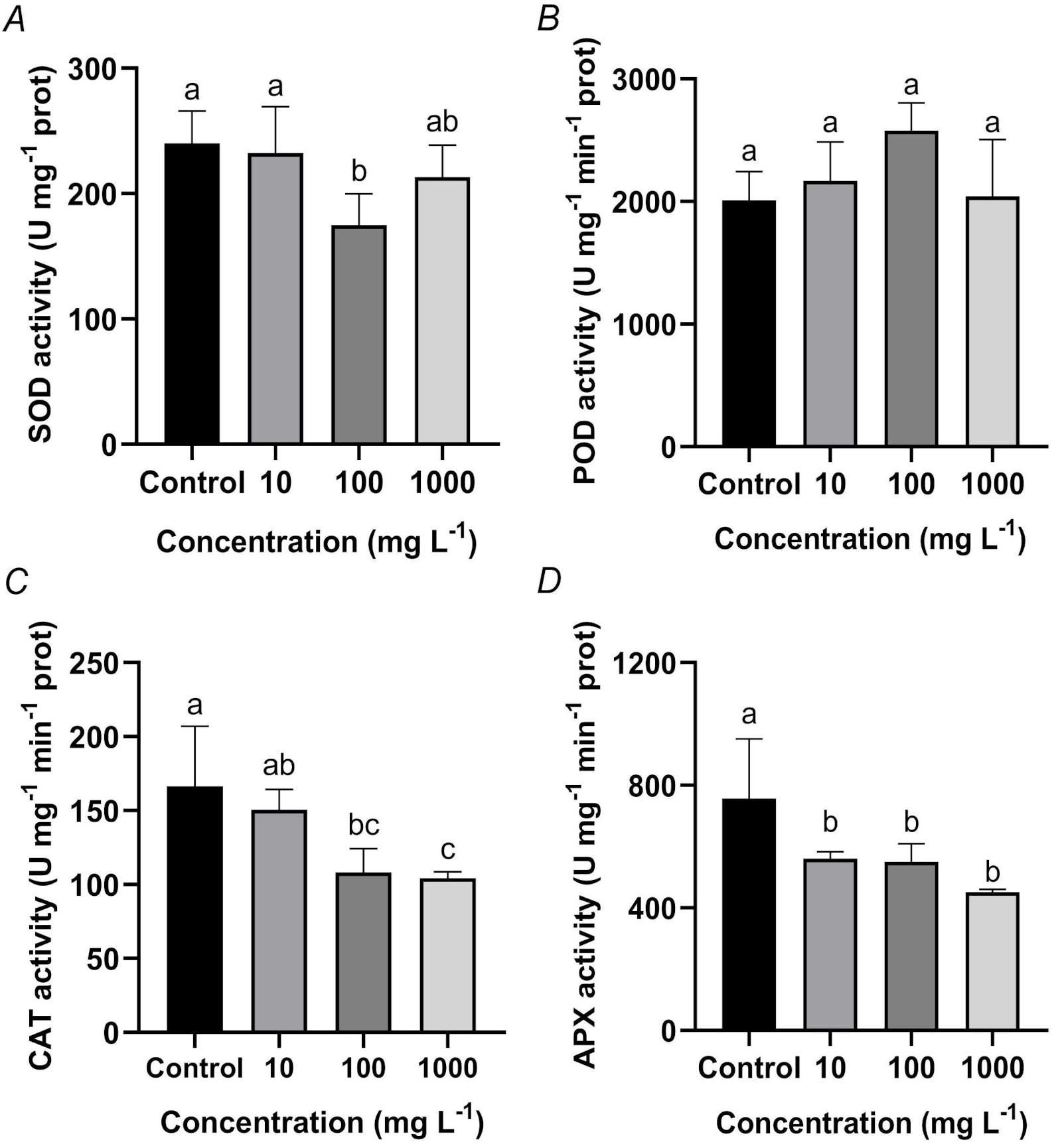
Fig. 6. Effects of a 10-day exposure to TWMs (10, 100, and 1 000 mg L-1) on the activities of root antioxidant enzymes in rice seedlings. (A) Superoxide dismutase (SOD). (B) Peroxidase (POD). (C) Catalase (CAT). (D) Ascorbate peroxidase (APX). Data are presented as mean ± SD (n = 3). Different letters indicate significant differences between the treatments (P < 0.05, Duncan's multiple test).
Discussion
Research on the ecological risks of microplastics has predominantly focused on common thermoplastics (Mateos-Cárdenas et al., 2021; Rozman et al., 2022). In contrast, the ecological risks of TWMs – a major category of thermosetting microplastics that account for over 50% of environmental microplastic pollution (Zhang et al., 2023) – remain poorly understood. Given their distinct material properties – particularly their persistent cross-linked structure and resistance to thermal chemical degradation (Baensch-Baltruschat et al., 2020) – as well as their potential species-specific toxic effects on key food crops such as rice, investigating TWMs is essential for a comprehensive assessment of overall microplastic risks. To facilitate a comprehensive assessment of microplastic risks in agricultural systems, the present study systematically evaluated the physiological effects of TWMs on rice seedlings by analyzing growth parameters, photosynthetic performance, and antioxidant system responses.
This study found that exposure to TWMs promoted the early growth of rice seedlings, with shoot height and root length ncreasing in a TWMs treatment-dependent manner (Fig. 1). This phenotypic promotion appears paradoxical to the commonly reported inhibitory effects of microplastics (Wang et al., 2023b). However, this phenomenon is consistent with the conclusion of concentration-dependent growth promotion of Chlorella vulgaris by TWMs observed by Yang et al. (2024), collectively confirming that specific microplastics can exert growth-stimulating effects within certain concentration ranges. Notably, the growth-promoting effect was accompanied by evident photosynthetic dysfunction and oxidative stress impairments (Fig. 2, Fig. 3, Fig. 4, Fig. 5, Fig. 6), implying a complex dose-response relationship of TWMs on rice seedlings. The resource allocation trade-off hypothesis may explain the contradictory phenomenon of growth promotion accompanied by physiological inhibition observed in our study: plants divert resources from maintaining photosynthetic efficiency and antioxidant defense capacity to prioritize the enhancement of shoot height and root length phenotypes, resulting in an unsustainable compensatory state that undermines long-term physiological resilience (Yamaji et al., 2003). Consequently, prolonged TWMs exposure is predicted to ultimately convert the initial growth stimulation into growth suppression (Zhao et al., 2024). This predicted transition from stimulation to suppression has been corroborated by a life-cycle study on mung bean exposed to TWMs (Azeem et al., 2024). That study demonstrated that TWMs adhered to root surfaces, reducing water and nutrient uptake, and led to significant yield reductions of 11, 28, and 52% at 0.01, 0.1, and 0.25% TWMs, respectively. These findings suggest that the initial compensatory growth observed in short-term exposures may ultimately translate into productivity losses over the complete life cycle.
Under TWMs exposure, the Y(II) of PSII in rice seedlings remained stable, indicating that the fundamental light-energy conversion capacity was not impaired (Fig. 2A) (Huo et al., 2025). However, the photochemical capacity of PSII, represented by qP (Gao et al., 2016), showed concentration-dependent variations. A steep decline in qP under low-concentration TWMs treatment (10 mg L-1) suggests a restriction in photochemical processes (Gebara et al., 2024), whereas the attenuated decrease in qP at high concentration (1 000 mg L-1) implies that PSII reaction centers may sustain functionality through potential compensatory mechanisms (Fig. 3A) (Havaux et al., 1991). In photosynthesis, excess light energy can be dissipated via regulated pathways, including NPQ and Y(NPQ), or through Y(NO) as non-light-induced non-photochemical fluorescence quenching loss (Shin et al., 2020; Qi et al., 2025). In the present study, the suppressed rise in NPQ and Y(NPQ) across all treatments, coupled with the elevated dissipation as Y(NO) (Fig. 3B-D), indicates a reduced ability to safely dissipate excess light energy as heat. This impairment in photoprotective capacity likely aggravates photoinhibition (Rocha and Melão, 2024).
RLCs provide critical insights into the physiological status of plant photosynthesis (Li et al., 2008). Under TWMs stress, the overall downward shift of the RLCs was specifically manifested as a significant decrease in the rETR (Fig. 2C). Simultaneously, the rETRmax at light saturation was also suppressed (Fig. 2B). Notably, despite the stable maintenance of Y(II), the rETR was significantly reduced (Fig. 2). Combined with the above results, it can be inferred that TWMs impair photosynthetic function in rice primarily by inhibiting the photosynthetic electron transport chain downstream of the PSII reaction center, rather than acting on the PSII reaction center itself. This inhibitory effect aligns with the findings of Wang et al. (2024), who reported that exposure to 500 mg L-1 tire-derived microplastics significantly suppressed both the RLCs and the rETRmax in the dinoflagellate Alexandrium pacificum. The underlying mechanism may be attributed to TWMs interfering with the electron transfer process within the PSII reaction center, specifically from plastoquinone QA to the QB site, thereby obstructing the stabilization of light-induced charge separation and subsequent electron transfer to the quinone pool (Wan et al., 2015). The specific impairment of photosynthetic electron transport capacity, not accompanied by any disruption to the basal photochemical function of PSII, constitutes an early diagnostic feature of photosynthetic stress (Todorenko et al., 2020). Collectively, the chlorophyll fluorescence data demonstrate that TWMs exposure, although not causing direct injury to the PSII reaction centers in rice seedlings, markedly inhibited their photoprotective regulatory ability and led to a severe decline in photosynthetic electron transport efficiency.
Plants respond to microplastic stress by regulating antioxidant systems, with these physiological changes serving as reliable indicators of their internal stress status (Dong et al., 2021). The results of this study demonstrate that TWMs exposure significantly enhanced soluble protein content in both roots and leaves of rice seedlings (Fig. 4). This response aligns with observations in lettuce under polystyrene microplastic (PS-MPs) stress reported by Lian et al. (2024), indicating a potential protective adaptation mechanism in which plants maintain physiological homeostasis by upregulating amino acid metabolism and protein synthesis. Regarding oxidative stress, TWMs treatment suppressed the activities of SOD and POD in leaves (Fig. 5A,B), while a coordinated inhibition of SOD, CAT, and APX activities was observed in roots (Fig. 6A,C,D). These findings indicate that TWMs exposure exceeds the tolerance threshold of rice seedlings, leading to a substantial impairment of their antioxidant defense system (Li and Wang, 2025). The disruptive impact of TWMs on the antioxidant system is consistent with recent findings by Wasnik et al. (2026), who reported that exposure to TWMs leachates inhibited antioxidant enzyme activities in both leaves and roots of mung bean. Their study showed that high concentrations of TWMs significantly elevated MDA levels and reduced SOD activity, indicating that leachate-induced reactive oxygen species (ROS) accumulation triggers severe lipid peroxidation and cellular damage. At the molecular level, Zhang et al. (2021) demonstrated through transcriptomic analysis that microplastics downregulate genes involved in regulating antioxidant enzyme activity in rice roots, providing a mechanistic basis for the observed enzyme inhibition. This coordinated suppression of multiple antioxidant enzymes indicates a collapse in systemic compensatory capacity, ultimately compromising ROS scavenging efficiency and potentially increasing susceptibility to oxidative stress (Xu et al., 2025).
In summary, this study demonstrates that TWMs exposure likely drives a resource allocation trade-off in rice seedlings. Resources are reallocated from maintaining photosynthetic efficiency and antioxidant defense to promote the growth of rice seedlings. Thus, prolonged TWMs exposure is highly likely to reverse the initial growth stimulation into growth suppression. Notably, this study elucidates the toxic mode of action of TWMs in crops and emphasizes that risk assessment must not rely solely on apparent growth metrics but must focus on functional physiological damage. It should be acknowledged that TWMs are considered relatively low-activity pollutants (Weyrauch et al., 2025), and a 10-day exposure period may not fully capture their chronic toxicity. However, the physiological changes observed in this study demonstrate that TWMs can induce significant acute stress responses during the early developmental stages of rice seedlings. Thus, the early stress signals identified in this study may serve as predictors of long-term impairment in rice. Nevertheless, long-term exposure studies spanning the entire life cycle are warranted to validate the predicted transition from growth stimulation to suppression and to comprehensively assess the ecological risks of TWMs in agricultural systems.
Conclusion
In summary, this study demonstrates that while TWMs promote the short-term growth of rice seedlings, they also concurrently suppress electron transport in PSII, as evidenced by a significant reduction in the rETRmax, disrupt the dynamic balance between qP and NPQ, and impair photoprotective processes by inhibiting light-induced non-photochemical fluorescence quenching [Y(NPQ)] while elevating non-light-induced non-photochemical fluorescence quenching loss [Y(NO)]. Additionally, TWMs directly inhibit the activities of key antioxidant enzymes in both leaves and roots of rice seedlings. These impairments collectively force plants into an unsustainable compensatory state, in which short-term growth is achieved at the cost of compromised photosynthetic efficiency and antioxidant defense capacity. Therefore, the observed TWM-induced growth promotion should be regarded as a physiological misleading phenomenon, and long-term exposure is anticipated to ultimately lead to growth suppression. This work systematically clarifies the integrated disruptive effects of TWMs on the photosynthetic and antioxidant systems of crops, thereby providing critical physiological evidence for refining the ecological risk assessment of TWMs in agricultural environments.
Download citation

References
- Azeem, I., Adeel, M., Shakoor, N. et al. (2024) Co-exposure to tire wear particles and nickel inhibits mung bean yield by reducing nutrient uptake. Environmental Science: Processes and Impacts, 26, 832-842.
 Go to original source...
Go to original source... - Baensch-Baltruschat, B., Kocher, B., Stock, F. & Reifferscheid, G. (2020) Tyre and road wear particles (TRWP) - A review of generation, properties, emissions, human health risk, ecotoxicity, and fate in the environment. Science of The Total Environment, 733, 137823. Go to original source...
- Batool, T., Ali, S., Seleiman, M.F. et al. (2020) Plant growth promoting rhizobacteria alleviates drought stress in potato in response to suppressive oxidative stress and antioxidant enzymes activities. Scientific Reports, 10, 16975. Go to original source...
- Bradford, M.M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 72, 248-254. Go to original source...
- Campanale, C., Galafassi, S., Savino, I. et al. (2022) Microplastics pollution in the terrestrial environments: Poorly known diffuse sources and implications for plants. Science of The Total Environment, 805, 150431. Go to original source...
- Chae, E., Jung, U. & Choi, S.-S. (2024) Types and concentrations of tire wear particles (TWPs) in road dust generated in slow lanes. Environmental Pollution, 346, 123670. Go to original source...
- Ding, J., Zhu, D., Wang, H.-T. et al. (2020) Dysbiosis in the gut microbiota of soil fauna explains the toxicity of tire tread particles. Environmental Science & Technology, 54, 7450-7460. Go to original source...
- Dong, Y., Song, Z., Liu, Y. & Gao, M. (2021) Polystyrene particles combined with di-butyl phthalate cause significant decrease in photosynthesis and red lettuce quality. Environmental Pollution, 278, 116871. Go to original source...
- Eisentraut, P., Dümichen, E., Ruhl, A.S. et al. (2018) Two birds with one stone - fast and simultaneous analysis of microplastics: microparticles derived from thermoplastics and tire wear. Environmental Science & Technology Letters, 5, 608-613. Go to original source...
- Galafassi, S., Nizzetto, L. & Volta, P. (2019) Plastic sources: A survey across scientific and grey literature for their inventory and relative contribution to microplastics pollution in natural environments, with an emphasis on surface water. Science of The Total Environment, 693, 133499. Go to original source...
- Gao, M., Qi, Y., Song, W. & Xu, H. (2016) Effects of di-n-butyl phthalate and di (2-ethylhexyl) phthalate on the growth, photosynthesis, and chlorophyll fluorescence of wheat seedlings. Chemosphere, 151, 76-83. Go to original source...
- Gebara, R.C., Abreu, C.B.d., Rocha, G.S. et al. (2024) Effects of ZnWO4 nanoparticles on growth, photosynthesis, and biochemical parameters of the green microalga Raphidocelis subcapitata. Chemosphere, 353, 141590. Go to original source...
- Gopinath, P.M., Saranya, V., Vijayakumar, S. et al. (2019) Assessment on interactive prospectives of nanoplastics with plasma proteins and the toxicological impacts of virgin, coronated and environmentally released-nanoplastics. Scientific Reports, 9, 8860. Go to original source...
- Goßmann, I., Halbach M. & Scholz-Böttcher, B.M. (2021) Car and truck tire wear particles in complex environmental samples - A quantitative comparison with "traditional" microplastic polymer mass loads. Science of The Total Environment, 773, 145667. Go to original source...
- Goswami, S. & Das, S. (2016) Copper phytoremediation potential of Calandula officinalis L. and the role of antioxidant enzymes in metal tolerance. Ecotoxicology and Environmental Safety, 126, 211-218. Go to original source...
- Havaux, M., Strasser R.J. & Greppin, H. (1991) A theoretical and experimental analysis of the qP and qN coefficients of chlorophyll fluorescence quenching and their relation to photochemical and nonphotochemical events. Photosynthesis Research, 27, 41-55. Go to original source...
- Huo, H., Jaffar, M.T., Zhang, J. & Shang, J. (2025) Calligonum caput-medusae seedlings adapt to drought stress through changing chlorophyll fluorescence parameters. Frontiers in Plant Science, 16, 1640412. Go to original source...
- Kavian, S., Safarzadeh, S. & Yasrebi, J. (2022) Zinc improves growth and antioxidant enzyme activity in Aloe vera plant under salt stress. South African Journal of Botany, 147, 1221-1229. Go to original source...
- Li, L., Luo, Y., Li, R. et al. (2020a) Effective uptake of submicrometre plastics by crop plants via a crack-entry mode. Nature Sustainability, 3, 929-937. Go to original source...
- Li, Q.-M., Liu, B.-B., Wu, Y. & Zou, Z.-R. (2008) Interactive effects of drought stresses and elevated CO2 concentration on photochemistry efficiency of cucumber seedlings. Journal of Integrative Plant Biology, 50, 1307-1317. Go to original source...
- Li, S.-H. & Wang, P. (2025) Overexpression of Cucurbita pepo CpVQ27 increased susceptibility of tobacco to powdery mildew. Physiological and Molecular Plant Pathology, 137, 102614. Go to original source...
- Li, Y., Shi, T., Li, X. et al. (2022) Inhaled tire-wear microplastic particles induced pulmonary fibrotic injury via epithelial cytoskeleton rearrangement. Environment International, 164, 107257. Go to original source...
- Li, Z., Li, R., Li, Q., Zhou, J. & Wang, G. (2020b) Physiological response of cucumber (Cucumis sativus L.) leaves to polystyrene nanoplastics pollution. Chemosphere, 255, 127041. Go to original source...
- Lian, J., Liu, W., Meng, L. et al. (2021) Foliar-applied polystyrene nanoplastics (PSNPs) reduce the growth and nutritional quality of lettuce (Lactuca sativa L.). Environmental Pollution, 280, 116978. Go to original source...
- Lian, Y., Shi, R., Liu, J. et al. (2024) Effects of polystyrene, polyethylene, and polypropylene microplastics on the soil-rhizosphere-plant system: Phytotoxicity, enzyme activity, and microbial community. Journal of Hazardous Materials, 465, 133417. Go to original source...
- Liu, G., Wang, S., Feng, F. et al. (2025) Risks of microplastics on germination and growth of pepper (Capsicum annuum L.) depending on the type, concentration, and particle size. Environmental Technology and Innovation, 40, 104385. Go to original source...
- Luo, Z., Zhou, X., Su, Y. et al. (2021) Environmental occurrence, fate, impact, and potential solution of tire microplastics: Similarities and differences with tire wear particles. Science of The Total Environment, 795, 148902. Go to original source...
- Mateos-Cárdenas, A., van Pelt, F.N.A.M., O'Halloran, J. & Jansen, M.A.K. (2021) Adsorption, uptake and toxicity of micro- and nanoplastics: Effects on terrestrial plants and aquatic macrophytes. Environmental Pollution, 284, 117183. Go to original source...
- Osman, D.M., Yuan, W., Nyaga, M.P., Monikh, F.A. & Yang, Y. (2025) Dual acute effects of tire microplastics and their leachates on Spirodela polyrhiza and phyllosphere bacteria. Water Research, 287, 124362. Go to original source...
- Putar, U., Turk, K., Jung, J., Kim, C. & Kalčíková, G. (2025) The dual impact of tire wear microplastics on the growth and ecological interactions of duckweed Lemna minor. Environmental Pollution, 368, 125681. Go to original source...
- Qi, Z., Xu, C., Tang, R. et al. (2025) Response of photosynthesis and chlorophyll fluorescence to nitrogen changes in rice with different nitrogen use efficiencies. Plants, 14, 1465. Go to original source...
- Ralph, P.J. & Gademann, R. (2005) Rapid light curves: A powerful tool to assess photosynthetic activity. Aquatic Botany, 82, 222-237. Go to original source...
- Rocha, G.S. & Melão, M.G.G. (2024) Does cobalt antagonize P limitation effects on photosynthetic parameters on the freshwater microalgae Raphidocelis subcapitata (Chlorophyceae), or does P limitation acclimation antagonize cobalt effects? More questions than answers. Environmental Pollution, 341, 122998. Go to original source...
- Rozman, U., Jemec Kokalj, A., Dolar, A., Drobne, D. & Kalčíková, G. (2022) Long-term interactions between microplastics and floating macrophyte Lemna minor: The potential for phytoremediation of microplastics in the aquatic environment. Science of The Total Environment, 831, 154866. Go to original source...
- Sathicq, M.B., Sabatino, R., Di Cesare, A. et al. (2022) PET particles raise microbiological concerns for human health while tyre wear microplastic particles potentially affect ecosystem services in waters. Journal of Hazardous Materials, 429, 128397. Go to original source...
- Shin, Y.K., Bhandari, S.R., Cho, M.C. & Lee, J.G. (2020) Evaluation of chlorophyll fluorescence parameters and proline content in tomato seedlings grown under different salt stress conditions. Horticulture, Environment, and Biotechnology, 61, 433-443. Go to original source...
- Todorenko, D., Timofeev, N., Kovalenko, I., Kukarskikh, G., Matorin, D. & Antal, T. (2020) Chromium effects on photosynthetic electron transport in pea (Pisum sativum L.). Planta, 251, 11. Go to original source...
- Wan, J., Guo, P., Peng, X. & Wen, K. (2015) Effect of erythromycin exposure on the growth, antioxidant system and photosynthesis of Microcystis flos-aquae. Journal of Hazardous Materials, 283, 778-786. Go to original source...
- Wang, B., Qiao, D., Wen, B. et al. (2024) Tire microplastic particles and warming inhibit physiological functions of the toxic microalga Alexandrium pacificum. Journal of Hazardous Materials, 480, 136087. Go to original source...
- Wang, Y., Bai, J., Wen, L. et al. (2023b) Phytotoxicity of microplastics to the floating plant Spirodela polyrhiza (L.): Plant functional traits and metabolomics. Environmental Pollution, 322, 121199. Go to original source...
- Wang, Y., Wu, Y., Pu, Q. et al. (2023a) Aquatic toxicity of tire microplastics on marine and freshwater organisms: An in silico approach. Chemosphere, 313, 137523. Go to original source...
- Wasnik, S., Cheng, S., Keang, K., Zhang, H. & Cross, J.S. (2026) Impact of tire particles and tire leachate contaminants on plant physiology and soil health: Case study in mung bean and tomato. NanoImpact, 41, 100601. Go to original source...
- Weyrauch, S., Seiwert, B., Voll, M. & Reemtsma, T. (2025) Environmental aging of tire and road wear particles and tire additives: a long-term field study. Environmental Science: Processes and Impacts, 27, 3498-3505. Go to original source...
- Wik, A. & Dave, G. (2009) Occurrence and effects of tire wear particles in the environment - A critical review and an initial risk assessment. Environmental Pollution, 157, 1-11. Go to original source...
- Xu, W., Xiao, Y., Xu, X. et al. (2025) Imidacloprid exacerbates the physiological and biochemical effects of rice (Oryza sativa L.) under salt stress: A systematic evaluation based on chlorophyll synthesis and oxidative stress. Plant Physiology and Biochemistry, 229, 110694. Go to original source...
- Yamaji, K., Julkunen-Tiitto, R., Rousi, M., Freiwald, V. & Oksanen, E. (2003) Ozone exposure over two growing seasons alters root-to-shoot ratio and chemical composition of birch (Betula pendula Roth). Global Change Biology, 9, 1363-1377. Go to original source...
- Yang, Y., Liu, J., Lu, H. et al. (2024) Effects of tire wear particle on growth, extracellular polymeric substance production and oxidation stress of algae Chlorella vulgaris: Performance and mechanism. Aquatic Toxicology, 276, 107118. Go to original source...
- Zhang, M., Yin, H., Tan, J. et al. (2023) A comprehensive review of tyre wear particles: Formation, measurements, properties, and influencing factors. Atmospheric Environment, 297, 119597. Go to original source...
- Zhang, Q., Zhao, M., Meng, F., Xiao, Y., Dai, W. & Luan, Y. (2021) Effect of polystyrene microplastics on rice seed germination and antioxidant enzyme activity. Toxics, 9, 179. Go to original source...
- Zhao, W., Deng, J., Wang, J., Ge, C. & Yang, H. (2024) Adverse effects of microplastics on the growth, photosynthesis, and astaxanthin synthesis of Haematococcus pluvialis. Science of The Total Environment, 954, 176427. Go to original source...
- Zuo, G., Mei, W., Feng, N. & Zheng, D. (2025) Photosynthetic performance index (PIabs) and malondialdehyde (MDA) content determine rice biomass under combined salt stress and prohexadione-calcium treatment. BMC Plant Biology, 25, 823. Go to original source...